



|
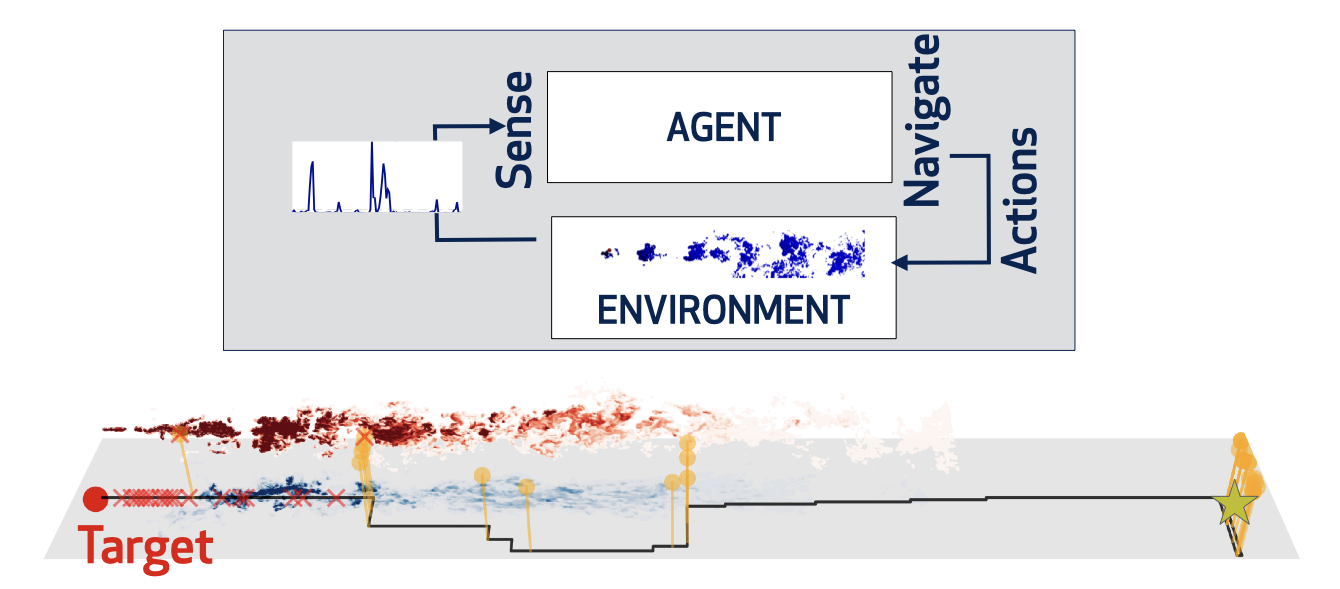
Organisms solve sophisticated olfactory problems even when odor cues are broken by turbulence. But (how) can an agent predict the position of a target using a short extract of the odor it releases? We develop supervised learning algorithms to demonstrate that measures of timing and intensity of odor detections enable olfactory predictions. However, to maintain predictive power, animals must rely on different measures as they move which -we predict- will be reflected in both behavior and neural representations of odor dynamics. The next question is: are the best predictors also the best features to support navigation? |
|
We develop reinforcement learning algorithms adapted to turbulent navigation, a paradigmatic example of decision making under uncertainty. In our recent work we use the framework of partially observable Markov Decision Processes to clarify why animals often alternate between sniffing the ground and the air while tracking scents. We are currently studying the foundations of these algorithms in the context of olfactory navigation to optimize their performance and extend the results to other multimodal sensory cues and a changing physical environment. |
|
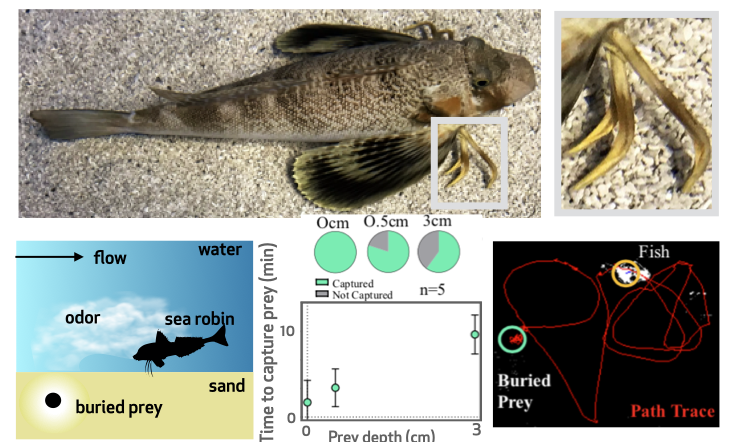
Fish have developed sophisticated sensory organs to perform behavior in the presence of turbulence. Although the algorithms implemented by fish during navigation remain largely obscure, a growing literature suggests that chemical and mechanical information are needed in tandem to explain behavior. Some fish even evolved novel organs to adapt to a particular niche. For example, besides the lateral line and the olfactory apparatus found in other fishes, sea robins evolved unique sensory organs that consist of six leg-like appendages used for walking and to dig prey buried beneath the seafloor. These leg-like structures are evolutionary novelties and respond to chemical, mechanical and proprioceptive stimuli to enable effective capture of buried preys. Searobins are so adept at finding prey that other fishes follow them to steal otherwise undetectable prey. The emergence of these novel sensory organs suggests that distinct algorithms are needed to find a target lying on the seafloor vs in the water column. We are interested into understanding the computational principles that shape the fascinating sensory biology of sea robins and underlie sensing and navigation in different environments. |

|
Sensory-guided navigation is one of the fundamental functions of the brain. A complete model of this behavior entails establishing the algorithms used by the brain to convert sensory cues into directional information as well as the neural circuit implementation of these algorithms. The mouse is a major model system in which these levels of understanding can be established using powerful genetic and neurophysiological tools. Although mice rely upon odors as vital navigational cues, little is understood regarding the algorithms and neural implementation for odor-guided navigation by mice. One main difficulty is that odors are dispersed by turbulent airflow into dynamic plumes that fluctuate dramatically in space an time. Measures of an odor even within a few centimeters of a mouse do not accurately report the odor arriving at its nose. We have demonstrated with experiments and theory that olfactory searches at close range are relatively easy because of the abundance of odor. But odor signals change as the searcher moves away from the source. We investigate how mice behavior as well as neural representations change depending on the dynamics of the underlying odor. |
|
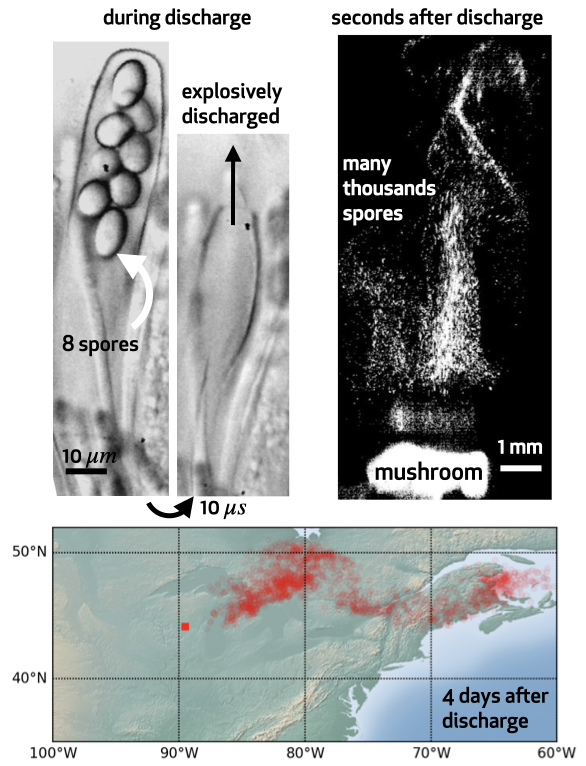
Microscopic spores allow higher fungi to disperse between ephemeral habitats. Spore range is an important factor in a species evolutionary success, since the probability of spore germination depends on the environment where it eventually lands. Although no range of dispersal can guarantee success, there is at least one important constraint that spores need to fullfill in order to germinate: they need not fall back on the parent fungus. This might seem quite an easy task, especially if we consider that spores are launched at enormous velocities that range from 1 m/s to even 20 m/s and corresponding accelerations of 10^5 g. However, at the micron scale air drag severely thwart particle dynamics and spores are decelerated to rest after travelling only few mm. This ballistic range is not large enough to cross the boundary layer of still air that surrounds the fruit body. For species that rely on wind to disperse their propagules this is a major obstacle, because spores that do not cross the boundary layer cannot be entrained by the dispersive air flows and eventually fall back on the parent fungus. We proposed that the microscopic mechanisms of spore discharge are optimized to ensure spores effectively disperse beyond the parent fungus. But the fate of a spore is ultimately dictated by atmospheric transport which is highly stochastic and beyond control of an individual fungus. We currently focus on spore dispersal in the atmosphere, to understand what strategies can be put in place to release spores at times that maximize their chances of survival. To this end, we use machine learning coupled to simulations and models of atmospheric transport to ask whether features of a particle trajectory in the atmosphere can be predicted by conditions at spore release. |

|
Bacterial biofilms are heterogeneous colonies of differentiated bacteria that live in association with surfaces and present a remarkable spatio-temporal organization. The formation of a mature biofilm begins from cell attachment to a surface and proliferation. External signals trigger an irreversible commitment to the surface. The hallmark of biofilm formation is the dramatic decrease in flagella mediated motility and simultaneous production of an extracellular polymeric matrix encasing the cells and holding the community together. In the Gram-positive soil bacterium Bacillus subtilis, most cells across the biofilm eventually give up their individual motility. However, a slower kind of surface translocation arises that is not mediated by flagella and enables the biofilm to colonize the surface and increase nutrient uptake. In order to spread the biofilm needs to overcome friction with the surface: I proposed that biofilm metabolism creates osmotic forces that enable biofilm spreading. What is their relation with the spatiotemporal patterns of gene expression? Are forces actively controlled by the community? |